
Closing the Microeukaryotic Gap in Modern Biosurveillance
Why environmental surveillance should look beyond viruses and bacteria
Vanessa Smilansky
June 2, 2026
What are microeukaryotes and why should we care about them?
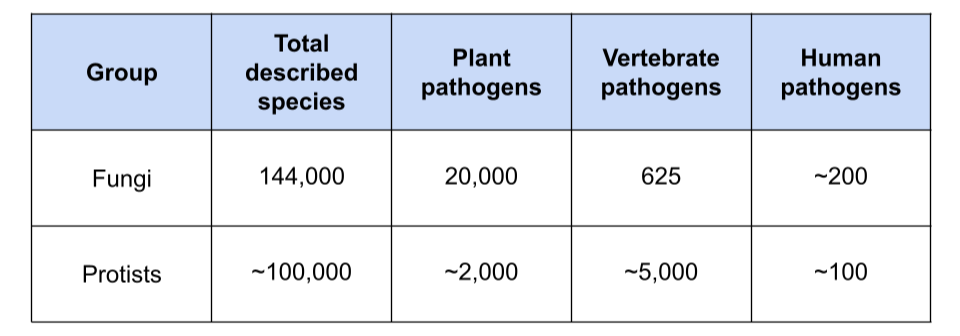
Viruses and bacteria dominate most discussions of microbes, yet they represent only part of the microbial world. Beyond them lies a diverse assemblage of microscopic single-celled eukaryotes that remains largely overlooked by modern biosurveillance efforts. These "microeukaryotes" are an important group of organisms that include fungi (filamentous fungi and yeasts), protists, and metazoan parasites (cestodes, nematodes, and helminths). Ubiquitous across terrestrial and aquatic environments, they play essential roles in ecosystem function, nutrient cycling, and food webs. At the same time, they are responsible for a substantial burden of disease in humans, animals, and plants (Table 1). This blog post focuses on pathogenic fungi and protists.
Table 1. Species diversity and pathogen distributions across fungi and protists. Fungi estimates are derived from Kew reports. Protist estimates are approximate and synthesized from multiple sources, reflecting the less complete taxonomic characterization of protist groups.
Fungi include some of the most significant microeukaryotic pathogens, ranging from devastating plant pathogens such as Magnaporthe oryzae (the agent of rice blast) to emerging human pathogens such as Candida auris. The chytrid fungus Batrachochytrium dendrobatidis has caused catastrophic population declines in hundreds of amphibian species and is regarded as one of the most destructive pathogens in terms of biodiversity loss. When not causing disease directly, fungi can also facilitate the transmission or persistence of other pathogens. Chytrid fungi in the genus Olpidium act as vectors for several plant necroviruses, facilitating disease spread in agriculture. Other fungi form biofilms that enhance the survival and colonization of bacterial pathogens, enabling their persistence in both clinical and environmental settings.
Protists likewise cause substantial disease across domains. One notable group is the alveolates, which include Plasmodium, the parasites that cause malaria, a disease responsible for more than 600,000 human deaths annually. Also within this group are the perkinsids, which comprise marine parasites of economically important bivalve molluscs, as well as a freshwater lineage that infects frog tadpoles (the focus of my PhD). Distinct from the alveolates, the kinetoplastids include Trypanosoma and Leishmania, parasites responsible for neglected tropical diseases (NTDs) affecting millions of people worldwide. Protists are also known vectors and reservoirs for pathogenic viruses and bacteria; for example, soil-borne Polymyxa protists transmit several plant mosaic viruses, while the bacteria causing Legionnaires’ disease can persist and replicate within free-living amoebae.
Many fungi and protists have complex life cycles that include highly resilient spore stages. Soil-borne pathogens such as the fungus Synchytrium endobioticum (the agent of potato wart disease) and the protist Plasmodiophora brassicae (the agent of clubroot) produce resting spores that can survive in soil for decades, making eradication difficult once established. Many species produce airborne reproductive spores that can travel hundreds of miles on wind currents, enabling rapid expansion into new regions. Some of these spores pose direct threats to human health. For example, Coccidioides fungi (the agents of Valley Fever) produce airborne spores that can cause severe respiratory disease when inhaled. Individuals with frequent environmental exposure, including workers in farming and construction, may be at particularly high risk.
Why do they matter for biosecurity now?
Climate change is creating new opportunities for pathogenic fungi and protists. Warmer water temperatures can increase transmission rates and shorten incubation periods for waterborne protists such as Cryptosporidium and Giardia, two of the most common causes of gastrointestinal disease. Frequent flooding and altered precipitation patterns can contaminate drinking water supplies, elevating the risk of disease outbreaks. Rising temperatures may also create evolutionary pressure for heat tolerance, allowing some fungal species to overcome the thermal barrier that normally protects mammals from infection. The multidrug-resistant yeast C. auris is hypothesized to be among the first human pathogens to emerge from this warming trend. As environmental conditions continue to shift, fungal and protistan pathogens may expand into new regions, encounter novel hosts, and generate outbreaks in previously unaffected populations.
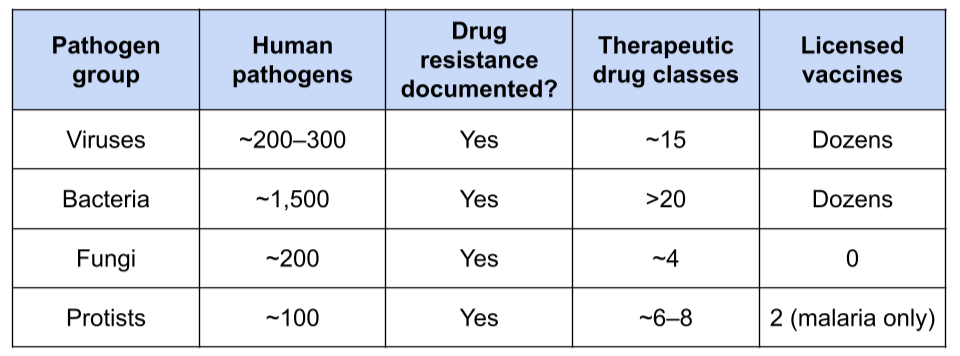
Rising antimicrobial resistance and limited countermeasures increase the risks posed by microeukaryotic pathogens. Drug-resistant fungi (e.g., C. auris, Aspergillus fumigatus) are growing public health concerns, while fungicide-resistant plant pathogens (e.g., Zymoseptoria tritici) threaten agricultural productivity. Resistance to antimalarial drugs has repeatedly evolved in Plasmodium falciparum, threatening decades of progress in malaria control. More broadly, the therapeutic and preventive toolbox for microeukaryotic infections remains limited (Table 2). Compared with antiviral and antibacterial drugs, there are relatively few antifungal and antiparasitic treatments, and no licensed vaccines currently exist for any human fungal disease. Although licensed malaria vaccines have recently become available, they remain the exception rather than the rule among most protistan diseases. As resistance continues to evolve, treatment options may become increasingly constrained.
Table 2. Therapeutic and vaccine countermeasures available for major pathogen groups. Values are approximate and intended to illustrate broad differences in countermeasure availability rather than provide definitive counts.
Microeukaryotes remain largely absent from modern biosurveillance systems. Most of these systems are optimized for viruses and bacteria, despite fungi and protists posing significant risks to public health, agriculture, and ecosystems. This oversight is especially concerning in the age of AI, as advances in synthetic biology, genome editing, and machine learning-guided protein design expand our ability to manipulate increasingly complex organisms. Although microeukaryotes are currently more difficult to engineer than viruses or bacteria due to their larger genomes, more complex cellular biology, and intricate life cycles, this barrier may not remain static. As biotechnology continues to advance, they may become more tractable and therefore more consequential from a biosecurity perspective. Closing the microeukaryotic gap will be essential for building a more complete picture of biological risk and strengthening early-warning surveillance.
What would a microeukaryote-aware biosurveillance system look like?
Effective surveillance of microeukaryotes requires an environmental perspective (Figure 1). While some important biothreats, such as C. auris, are primarily associated with healthcare settings, many fungi and protists persist and spread through environmental reservoirs long before they are detected in human, animal, or plant hosts. Consequently, surveillance should prioritize the air, water, and soils where these organisms circulate. In practice, this could include monitoring drinking water networks, agricultural systems, and human–wildlife interfaces where pathogen spillover is most likely to occur. Complementary post-exposure surveillance could target environments where biological signals accumulate following infection, such as municipal wastewater and livestock manure lagoons. Together, these approaches may provide earlier warning of emerging fungal and protistan threats and help protect public health, food security, and biodiversity.
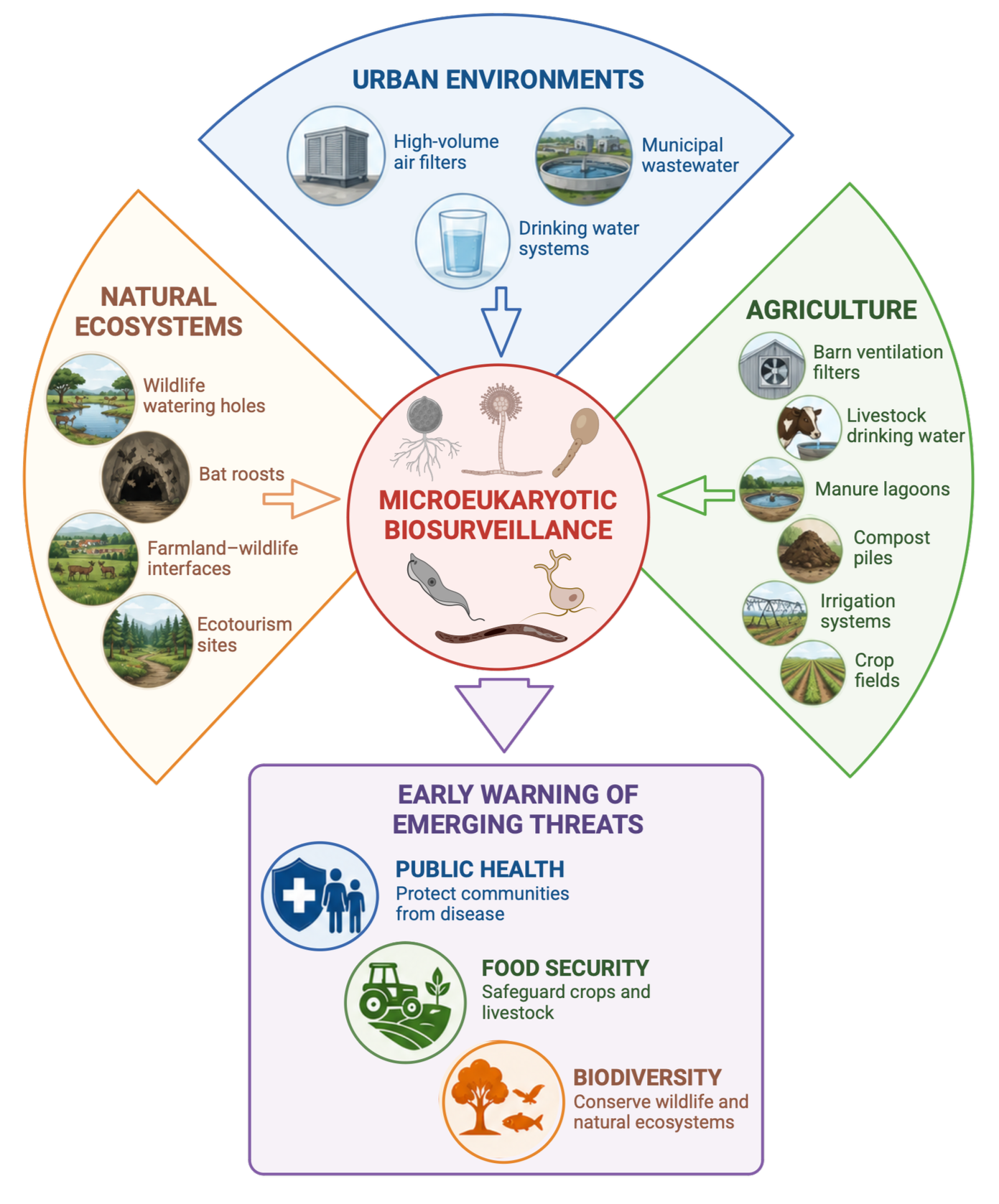
Figure 1. One Health framework for environmental biosurveillance of microeukaryotes. Created in BioRender. Smilansky, V. (2026) https://BioRender.com/02o6f3l
Metagenomic pipelines should be adapted to better capture microeukaryotes. Because metagenomic sequencing does not depend on predefined targets, it is well suited for detecting novel, engineered, or highly divergent organisms. However, targeted methods may remain preferable for monitoring specific known threats, where higher sensitivity, lower cost, and more streamlined workflows are advantageous. Existing metagenomic pipelines are typically optimized for viruses and bacteria and can fail to recover microeukaryotes during sample processing and sequencing, while bioinformatic approaches struggle to assemble and classify their large and often repetitive genomes.
To address these limitations, various steps in the pipeline–including size selection, nucleic acid extraction, library preparation, sequencing, and bioinformatic analysis–should be optimized for the recovery, detection, and classification of informative microeukaryotic sequences. For example, mRNA can be used instead of, or in addition to, DNA to identify metabolically active organisms, helping distinguish viable pathogens from dormant spores, dead cells, and residual eDNA. Long-read sequencing could be prioritized over short-read sequencing to better handle repetitive regions during genome assembly. Taxonomic classification is challenging because most microeukaryotic diversity remains absent from reference databases; however, this limitation should improve over time as more species are characterized through efforts to map the eukaryome, such as the Earth BioGenome Project. At the same time, emerging "function-first" bioinformatic approaches may reduce reliance on taxonomy by identifying biosecurity-relevant features directly from raw metagenomic data. Finally, reporting frameworks will be needed to translate complex metagenomic data into actionable information. Taken together, these pipelines could support the identification of known and novel microeukaryotic pathogens, antimicrobial resistance markers, genetic modifications, and other signals that warrant further investigation or intervention.